Levende liv
(Utvalgte deler fra Kap. 9-10; i 'Undeniable' av Douglas Axe, Harper One)
 I boka 'Undeniable' behandler Douglas Axe konflikten mellom en design intuisjon og rådende konsensus-syn i naturvitenskapen, om livets opprinnelse og utvikling. Han benytter argumenter fra en allmenn, 'folkelig vitenskap', ved støtte av egen forskning om proteiners utforming og virkemåte. Men han går også videre, i det han ønsker å finne svar på 'det store spørsmålet': 'Hva eller hvem skylder vi vår eksistens?' Selv om knapt noe menneske våkner opp om morgenen og takker naturlig seleksjon eller blinde repetisjoner for sin eksistens, så har vi fra ID-hold bare et vagt svar på det store spørsmålet. Men om vi ikke er avkom fra noen utilsiktet årsak i det hele, så blir hensikt en nøkkel-ingrediens i vårt opphav. Og mange fra ID-hold ønsker kanskje å la det forbli med det?
I boka 'Undeniable' behandler Douglas Axe konflikten mellom en design intuisjon og rådende konsensus-syn i naturvitenskapen, om livets opprinnelse og utvikling. Han benytter argumenter fra en allmenn, 'folkelig vitenskap', ved støtte av egen forskning om proteiners utforming og virkemåte. Men han går også videre, i det han ønsker å finne svar på 'det store spørsmålet': 'Hva eller hvem skylder vi vår eksistens?' Selv om knapt noe menneske våkner opp om morgenen og takker naturlig seleksjon eller blinde repetisjoner for sin eksistens, så har vi fra ID-hold bare et vagt svar på det store spørsmålet. Men om vi ikke er avkom fra noen utilsiktet årsak i det hele, så blir hensikt en nøkkel-ingrediens i vårt opphav. Og mange fra ID-hold ønsker kanskje å la det forbli med det?
I tilfellet Douglas Axe så er han blitt overbevist av ateisten Thomas Nagel (1), at vi skulle prøve å komme videre. Ateisten Nagel forsøker å 'forklare tilblivelsen av liv, bevissthet, fornuft og kunnskap verken som tilfeldige bivirkninger av fysiske naturlover, eller som resultat av hensiktsstyrt inngripen i naturen fra utsiden, men som en ikke-overraskende, om ikke uunngåelig konsekvens av den orden som styrer naturen fra innsiden." Selv om Nagel er ateist, så er han altså enig med Intelligent Design (ID) i at utilsiktede årsaker ikke danner grunnlag for livet.
Bilde 1. Hvordan barn kan møte evolusjonsteorien
Så selv om Nagels støtte er bra opp til ett punkt, at man må går for enten en ubevisst eller personlig årsak bakom tilværet, så ville det være et uklart standpunkt å ende opp med det første valget i så måte. Derfor trenger vi å fortsette med sikte på en klarere forståelse av denne ikke-utilsiktede kilden, som vi skylder vår eksistens til. Vi vil prøve å benytte sunt folkevett, og benytte en analyse av hva nyvinninger eller oppfinnelser, innebærer. Vi skal også se på noen vanlige innvendinger mot design-intuisjonen som har blitt rettet mot ID.

Vi vil gå for det synet at det som ser ut som det er resultat av ett geni, er frukt av ett geni. Den universelle design-hypotesen påstår at det er slik, og alt i vår daglige erfaring bekrefter den påstanden. Oppfinnelser er ting som krever dyktighet, og ting som krever dyktighet må oppnås gjennom dyktighet. Oppfinnere søker av og til etter bedre måter å gjøre ting på, men de søker ikke i blinde. Oppfinnelser handler tross alt om 'mentale lyspærer' som viser oss ting klart for første gang. Det ligner ikke på målløs famling i mørket, noe som karakteriserer et blindt søk.
Bilde 2. Petter Smart's 'lille-hjelper'
Hvordan mennesker finner opp
Uten å forutsette at alle oppfinnelser må skje på det viset menneskelige oppfinnelser gjøres, så vil det hjelpe å tenke på hvordan vi mennesker finner opp. Forfatteren innser at flesteparten av oss ikke oppfatter seg selv som oppfinnere, noe mer enn vi oppfatter oss som vitenskapsfolk. Men det kan skyldes at vi undervurderer betydningen av det vi gjør av rutine, med letthet. Om vi kikke nærmere etter hva oppfinnelser er, ser vi at oppfinnelser er en vesentlig del av det å være ett menneske. For å hjelpe oss å komme dit, har forfatteren brutt ned prosessen om menneskelig oppfinnelse, ned i 3 stadier. Det første stadiet av disse, er det mentale stadiet. Det er der den opprinnelige idéen til oppfinnelsen blir utviklet til en detaljert plan, klar til å implementeres. Den begrepsmessige hoved-idéen må alltid brytes ned i mindre idéer, som kanskje igjen må brytes ned i mindre, før iverksettingen kan starte. Det andre stadiet er hvor den resulterende, begrepsmessige planen, må brukes til å konstruere en fysisk gjenstand.
Det mentale arbeid på dette stadiet, er mer praktisk enn det var i første stadium. Den fullt unnfangede planen blir iverksatt på fysisk materiale. Dette krever evne til både å løse alle detaljspørsmål som oppstår når komplekse planer implementeres for første gang. Når dette kalles den metodiske fasen, så er det for å omfatte både bevisst, hensiktsstyrt aktivitet, og at denne tar behørig hensyn til egenskapene ved fysiske objekter og materialer som manipuleres. Poenget med første fase, var å danne idéer ovenfra-og-ned, mens i den andre fasen, er det å forme en fysisk enhet fra bunnen og oppover, i det en beveger seg fra råmateriale til en fungerende prototype.

Om alt fungerer greit, så vil den overordnede idéen klart komme til uttrykk i 3. stadium, i det den ferdige enheten gjør det den er tenkt å gjøre. Det kan sammenlignes med f.eks. en basseng-robot. Alle som ser en slik enhet i aktivitet, vil etter hvert komme på idéen at den faktisk gjør rent i bassenget. I det vi observerer fysisk aktivitet, slutter vi at tidligere bevisst aktivitet produserte den spesielle formen for fysisk aktivitet, nemlig aktiviteten til en fysisk helhet. Ved siden av å bevitne oppfinnelsen i virksomhet, slutter vi at den ble konstruert ifølge en unnfanget plan.
Fig 1. Menneskelige oppfinnelser gjennomløper 3 stadier
Å ta oppfinnelser til et helt nytt nivå
Uansett hvor evnerikt mennesker har høstet fra naturens regulariteter, ved å forme elementene til gjenstander som smart-telefoner og rom-teleskop, så kommer vi vanskelig unna aktualiseringen av at noen har overgått oss. Den travle edderkoppen, den heroiske laksen, den kraftfulle spekkhoggeren, ja faktisk alt levende som omgir oss, demonstrerer at fysiske materialer og prosesser kan bli satt sammen mye mer elegant enn vi noensinne har gjort. Forfatteren har selv vært entusiastisk tekno-nerd hele livet, så han sier ikke dette for å nedvurdere menneskelige oppfinnelser, men for å minne om at livet inneholder en kategori som udiskutabelt befinner seg ovenfor disse.
Blant de mer avanserte produkter av menneskelig teknologi, er et soldrevet undervannsfartøy, kalt 'Tavros 2'. Denne ble designet for å drive månedslange oppdrag i Mexico-gulfen, og driftes av universitetet i Sør-Florida. Tavros 2 måler og rapporterer dybde og temperatur, men det som gjør denne farkosten sofistikert, er at den opererer autonomt, under kontroll av sin interne komputer. Tavros 2 er programmert til å stige til overflaten når den trenger energi-påfylling. Etter dette går den tilbake til opprinnelig plassering og fortsetter data-innsamling. Så GPS-navigering, og tweeting av data, samt dens robot-egenskaper, er av dens tekniske ferdigheter (2).
Om en skulle sammenligne Tavros 2 med noe levende av noenlunde samme størrelse, kunne en f.eks. tenke på delfiner. Men ikke før en slik sammenligning er påbegynt, innser vi at de to er meget forskjellige. Som alle roboter, gjør Tavros 2 alt den er programmert til å gjøre, mens delfiner synes å gjøre alt de selv ønsker å gjøre. Den ene er en fysisk maskin, mens den andre er, etter ytre å dømme, noe større enn dette. Denne dype forskjellen kommer Axe tilbake til senere, men nå skal vi befatte oss med de fysiske aspektene ved levende liv, aspekter som kan ha noen likhetstrekk med maskiner, av det mest bemerkelsesverdige slaget. Vi skal se at livets maskineri fremviser funksjonell konsistens utover menneskelig forståelse, for ikke å snakke om menneskelig oppfinnelser.
High-Tech 'dam-slagg'
Vi skal starte med en 'enkel' livsform, en laverestående vann-mikrobe, kalt cyanobakterie. Selv om cyanobakterien er en enkelt-celle organisme, så kan individene til noen arter, knytte seg til hverandre og danne lange tråder, som fletter seg sammen til store kolonier i stillestående eller sakte-flytende vann. De er bokstavelig talt jordens 'dam-slagg'. Selv om de okkuperer en lav posisjon i livets store system, så er cyanobakterien lysår foran Tavros 2 i forhold til teknisk raffinement. La oss sammenlikne litt: Én kjent likhet er at Tavros 2 og cyanobakterien begge er solar-drevne. Men om vi undersøker den egenskapen i større detalj, finner vi at de to ikke er sammenlignbare. Den ikke-levende maskinen trenger en sol-oppsamler på størrelse med ett kaffe-bord, mens den levende klarer seg godt med én som er en billionte-del av den størrelsen. Og mens den ikke-levende bare har ett trick for å samle solenergi: å stige til overflaten, så er den levende i stand til mye mer. Trådaktige cyanobakterier kontrollerer dybden i forhold til sollyset, men de er også i stand til å koordinere komplekse glidebevegelser og oscillerende bevegelser, så hele kolonien vender seg mot sollyset.
Kontrasten blir enda tydeligere om en ser på framstillings-muligheter. Tavros 2 har ingen, mens hver cyanobakterie innehar et helt framstillingskart, innenfor sine mikroskopiske vegger. Som energikilde for alle funksjoner i denne planten, er prosessen kjent som fotosyntese, overføring av lysenergi til kjemisk energi. Mye av denne kjemiske energien, blir benyttet til å danne sukker-molekyler av CO2 og vann, som gir oksygen (O2) som et biprodukt. Sukker, er energirikt, slik at celler kan forbrenne det for å fungere. Alternativt fungerer sukker som en mangesidig karbon-forbindelse, som celler kan benytte for å danne den store variasjon av andre karbon-rike molekyler, som trengs i levende liv.
Selv om fotosyntese foregår rundt omkring oss i naturen, så er den på en måte motsatt av naturlige prosesser. Mer enn noen menneskelige oppfinnelser, så er fotosyntesen en oppfinnsom utnyttelse av naturlige regelmessigheter i naturen. Samtidig radikalt annerledes fra noen av disse regelmessighetene i seg selv. Fotosyntese kan betraktes som den motsatte prosessen av å forbrenne drivstoff. Å forbrenne noe, er en meget naturlig prosess, mens å 're-forbrenne' (unburn) noe, ikke er det. Med bare en gnist til å starte den, så fortærer oksygen drivstoff-molekyler, lik sukker, og danner CO2 og vann i dampform. Ved å gjøre det -motsatte,  så fortjener fotosyntesen plass som én av de fine oppfinnelser, lik air-condition, som høster naturlige regelmessigheter for å arbeide mot dem. Og av disse to oppfinnelsene, så er fotosyntesen absolutt den beste.
så fortjener fotosyntesen plass som én av de fine oppfinnelser, lik air-condition, som høster naturlige regelmessigheter for å arbeide mot dem. Og av disse to oppfinnelsene, så er fotosyntesen absolutt den beste.
Fig 2. Dele-liste for oppbygning av cyanobakterien
For å få fram virkemåten uten å måtte tvære det ut, går vi til indre funksjonalitet, for å se hvordan alle delene er arrangert på innsiden. Om vi ser på fotosystem I, en av hovedkomponentene i fotosyntese-funksjonaliteten til cyanobakterien, har den en inventarliste på 12 protein-deler og 6 mindre kofaktorer. Én av de mest brukte er klorofyll, som er benyttet 288 ganger for å danne hele foto-syntese systemet. Disse essensielle kofaktorene holdes presis på plass av et stort protein-rammeverk (Fig 2). Det komplette fotosystem I, vist på bunnen av Fig. 3, har 417 deler, som er presist posisjonert for at hele enheten skal utføre sin funksjon, med å samle fotoner fra sola, og omdanne lys-energien til kjemisk energi. Etter forfatterens beregning er det ca. 3 dusin gener, i cyanobakteriens genom, som er dedikert til å bygge denne sammensetningen. Ett dusin for protein-kodende komponenter, og mer enn to dusin for å kode enzymer som trengs til å framstille kofaktorene. I molekylære termer, er dette en massiv sammensetning, men med en diameter på bare 22 milliardte-dels meter, så er det plass til ca 15 millioner av disse på området til ett pixel på en iPhone Retina skjerm.
 For de som er interessert i hvordan antenne-systemet eller elektron-overføringssystemet virker, så er det gode ressurser tilgjengelig online. Men for å bli overbevist om at fotosystem I er en nano-teknologisk oppfinnelse av høy-teknologisk standard, så kan en i utgangspunktet betrakte diagram som illustrerer dem (Fig. 3 og 4).
For de som er interessert i hvordan antenne-systemet eller elektron-overføringssystemet virker, så er det gode ressurser tilgjengelig online. Men for å bli overbevist om at fotosystem I er en nano-teknologisk oppfinnelse av høy-teknologisk standard, så kan en i utgangspunktet betrakte diagram som illustrerer dem (Fig. 3 og 4).
Fig 3 cyanobakteriens fotosystem I, med to av dens viktige komponenter (antenne-system og elektron overførings-system)
Det selvsamme faktum at termene 'elektron overførings-system' og 'antenne-system' blir benyttet av vitenskapsfolk som studerer fotosyntese I, viser at denne fotosyntese I involverer flere underfunksjoner, inkludert overføring av elektroner og oppsamling av fotoner, via en antenne. Om du ønsker å dukke ned i den vitenskapelige litteraturen, finner du en rekke andre termer, som: 'dokking-system, primær elektron-donor, initiell elektron-mottaker, og sluknings-karotener'. Selv om de fleste ikke vet hva disse termene innebærer, så ser vi alle at høynivå-systemet til fotosyntese I, avhenger av et omfattende hierarki av lavere funksjoner. Det er et såkalt eks. på koherens i hierarkiske-funksjoner, som tydeliggjøres av den ørlille skalaen den er implementert på. For noen av oss fungerer dette mønsteret som signaturen av en oppfinnelse, med en hensikt. Selv om fotosyntese I er kompleks, er den bare én av mange komponenter som utgjør hele fotosyntese-systemet. En anelse av hvor komplekst systemet er, kan en se i Fig. 4. Figuren er arrangert i en hierarkisk struktur, der cyano-cellen vises som et tverrsnitts-bilde tatt med elektron-mikroskop. Under dette er fotosyntese systemet, som selv om det vises alene, bare er én av mange systemer som trengs for å støtte toppnivå-funksjonen av levende liv, lik cyanobakteriens celle.
På nivået under, så er fotosyntese systemet vist  sammensatt av to komponenter: thylakoid-membranen og CO2-konsentrasjon/reaktiverings-systemet. Det første er ansvarlig for å samle lysenergi og omdanne det til kjemisk energi. Det andre for å benytte dette som kjemisk energi til å 're-forbrenne' CO2. Hovedstrukturene til hver av dem er skjelnbare i toppbildet. De konsentriske bandene som sees rundt cellens periferi, er lagene av lys-oppsamlende thylakoid-membran. De mørke flekkene inni cella, er karboksyl-forbrenningsovner, der 're-forbrenningen' finner sted. Alle disse funksjonene krever eksplisitt teknisk funksjonalitet. F.eks. danner thylakoid-membranen avdelinger, som er så godt forseglet, at selv et lite proton (H+) ikke kan passere barrieren, uten å passere gjennom en sofistikert protein-kanal, som systematisk flytter den fra den ene siden til den andre. Noen av disse kanalene (fotosystem II og cytokrom b6f-komplekset) fungerer som små pumper, som flytter protoner fra 'lavtrykks' til 'høytrykks'-siden av seksjonen. En annen (ATP-syntase) fungerer som en turbin, og ekstraherer energi, ved å tillate protoner å gå motsatt vei.
sammensatt av to komponenter: thylakoid-membranen og CO2-konsentrasjon/reaktiverings-systemet. Det første er ansvarlig for å samle lysenergi og omdanne det til kjemisk energi. Det andre for å benytte dette som kjemisk energi til å 're-forbrenne' CO2. Hovedstrukturene til hver av dem er skjelnbare i toppbildet. De konsentriske bandene som sees rundt cellens periferi, er lagene av lys-oppsamlende thylakoid-membran. De mørke flekkene inni cella, er karboksyl-forbrenningsovner, der 're-forbrenningen' finner sted. Alle disse funksjonene krever eksplisitt teknisk funksjonalitet. F.eks. danner thylakoid-membranen avdelinger, som er så godt forseglet, at selv et lite proton (H+) ikke kan passere barrieren, uten å passere gjennom en sofistikert protein-kanal, som systematisk flytter den fra den ene siden til den andre. Noen av disse kanalene (fotosystem II og cytokrom b6f-komplekset) fungerer som små pumper, som flytter protoner fra 'lavtrykks' til 'høytrykks'-siden av seksjonen. En annen (ATP-syntase) fungerer som en turbin, og ekstraherer energi, ved å tillate protoner å gå motsatt vei.
Dette er bare et lite øyeblikksbilde av kompleksiteten til fotosyntesen. Volum har blitt skrevet om dette emnet. Og så forbløffende som den funksjonelle sammenhengen i Fig. 4 er, så blir det enda mer forbløffende når vi studerer det høyeste nivå i hierarkiet, der de mange funksjonene forenes i én hensikt. Fra dette gunstige toppnivå-punktet, ser vi at fotosyntese, til tross for alle dens forbløffende spissfindigheter, bare er én av mange hovedfunksjoner som trengs for at cyanobakterien skal fullføre sin hensikt, å være cyanobakterie. Sett gjennom denne linsen, så er fotosyntese én av de utsøkte oppfinnelser som tjener et høyere formål så godt, at det gjør seg selv nesten usynlig. Ut av alle de slående komplekse molekylene som sammenknyttes i overraskende imponerende systemer og superstrukturer, som trengs for å danne en levende cyanobakterie.
Fig. 4. Oppbygning til en cyanobakterie
Som konklusjon i sammenlikningen med Tavros 2, så får vi inntrykk av at den ikke er mer verdig å sammenlikne med en cyanobakterie, enn en høyerestående delfin. Det var tross alt med dyktig hjelp av tusenvis av mennesker ved hundrevis av industribedrifter av ulike slag, at Tavros 2 ble bygget. Med all mulig respekt, gjør denne industri-roboten lite i forhold til all menneskelig anstrengelse som er benyttet for lage den. Kontrasten i forhold til cyanobakterien kan nesten ikke være større. Med enestående genialitet forsynte oppfinneren av cyanobakterien den med evnen til å tilvirke cyanobakterier. Etter at den første cyanobakterien var tilvirket, hvordan enn det skjedde, så ble resten av cyanobakterier dannet av seg selv fra luft, solskinn og vann.
Koherens i steroider
 Cyanobakterien viser seg å være en imponerende helhet, ved å fullføre et tilsvarende imponerende helhetlig prosjekt: tilvirkningen av cyanobakterier -med overbevisende letthet. Om det er tilfelle, så kan knapt den hardt arbeidende edderkoppen eller den heroiske laksen innebære noe mindre. Cyanobakterien er bare begynnelsen. For å vise den elegante kompleksiteten som ligger under kjente aspekter ved høyerestående liv, så har forfatteren sporet en gren av synssystemet hos pattedyr, fra toppen av det komplekse, hierarkiske systemet som støtter syn, uten å måtte forstå det. Hvordan vi enn velger å representere dette systemet, hvor fint vi enn deler det opp, så er hierarkiet i seg selv meget reelt og imponerende. Og enda har vi bare krafset i overflaten, både hva bredde og dybde angår. Sannheten er at levende vesener er funksjonelt sammenhengende, på mye mer dyptgående vis enn hva menneskelige oppfinnelser er. Alt i en laks er på et mye mer diskriminerende vis, knyttet til toppnivå-hensikten: å være en laks. Kroppen er levende og trives, om alle deler fungerer, eller den er død og forråtnende om de ikke gjør det.
Cyanobakterien viser seg å være en imponerende helhet, ved å fullføre et tilsvarende imponerende helhetlig prosjekt: tilvirkningen av cyanobakterier -med overbevisende letthet. Om det er tilfelle, så kan knapt den hardt arbeidende edderkoppen eller den heroiske laksen innebære noe mindre. Cyanobakterien er bare begynnelsen. For å vise den elegante kompleksiteten som ligger under kjente aspekter ved høyerestående liv, så har forfatteren sporet en gren av synssystemet hos pattedyr, fra toppen av det komplekse, hierarkiske systemet som støtter syn, uten å måtte forstå det. Hvordan vi enn velger å representere dette systemet, hvor fint vi enn deler det opp, så er hierarkiet i seg selv meget reelt og imponerende. Og enda har vi bare krafset i overflaten, både hva bredde og dybde angår. Sannheten er at levende vesener er funksjonelt sammenhengende, på mye mer dyptgående vis enn hva menneskelige oppfinnelser er. Alt i en laks er på et mye mer diskriminerende vis, knyttet til toppnivå-hensikten: å være en laks. Kroppen er levende og trives, om alle deler fungerer, eller den er død og forråtnende om de ikke gjør det.
Fig 5. Komplett visuelt system
Menneskelige oppfinnelser er på langt nær så forenet i sin virkemåte, enten det er biler, smart-telefoner eller ulike slag roboter. De feiler om en komponent feiler, men i de fleste tilfelle blir ikke øvrige komponenter påvirket av den feilen. Mennesker finner (foreløpig) ikke opp så mange 'alt-eller ingenting-helheter'. I stedet finner vi opp del for del, og setter dem så sammen til helheter. Hver del testes uavhengig, ifølge egne spesifikasjoner. Livet fungerer annerledes, på nesten utrolig vis spiller levende 'oppfinnelser' en nøkkelrolle i å bygge og vedlikeholde seg selv, med alle sine deler formet og knyttet sammen i en helhet. Hver celle i kroppen styrker kroppen, og blir vedlikeholdt av kroppen.
Å få aminosyrer til å virke sammen
Tidligere har Douglas Axe vist via laboratorie-eksperimenter at Darwins molekylære tilbereder, naturlig seleksjon, ikke er i stand til å finne opp nye proteiner. Her har vi sett hvor dypt funksjonelle sammenhenger går gjennom biologiske systemer, oppbygd av proteiner (se Fig. 5). Disse to kan bli oppfattet som to separate problemer, i forhold til blinde evolusjonære søk. i) Problemet med å finne nye proteiner og ii) Problemet med hjelpsomme oppfinnelser, som benytter proteiner. Faktisk dreier begge problemene seg om det samme: umuligheten i å finne nødvendig funksjonell sammenheng ved blinde søk, fordi proteiner som molekylære oppfinnelser, utøver imponerende funksjonell sammenheng i seg selv.
Fig 6 hjelper oss å forstå hva funksjonell sammenheng innebærer i kontekst av en enkelt protein-kjede. Verdien av 'sløyfe-diagrammet' vist på venstre side, er at vi kan se hvor kjeden dannes enten på de to regulære formingene, som karakteriserer alle proteinfoldinger: enten alpha-helikser (vist som koiler) eller beta-tvinninger (vist som piler). Men den visuelle klarheten skjer på bekostning av overforenkling, som den mer fysisk nøyaktige 'kvist-representasjonen' til høyre viser. Mellom kvistene kan vi stedvis skimte en tagget versjon av den elegante stien til venstre i figuren, men vi ser også noe som minner om en masse forstyrrende 'rør', av tilføyelser som stikker ut i alle retninger. Tro det eller ikke, den funksjonelle sammenhengen til dette proteinet ligger innenfor hva dette komplekse 'røret' bestemmer, og det samme gjelder for alle foldede proteiner.
De ulike vedhengene er det som karakteriserer de ulike aminosyrene. Det som for oss ser ut som ett rot, er i virkeligheten et fortreffelig arrangement av aminosyre-vedheng, langs hele proteinet som styrer det som ellers  ville vært en lang kjede, i det å forme en stabil 3-dimensjonal struktur. Arrangert sekvensielt, på det viset, så passer vedhengene mer komfortabelt i deres foldede form, enn de ville gjort ved å flyte ustrukturert omkring i den cellulære væsken, slik som en tilfeldig sekvens av aminosyrer gjør. Uten deres fint-tilpassede formasjoner, kunne ikke livets proteiner utføre sine vitale funksjoner (4). I ett eksperiment prosjekt, satte Axe seg fore å finne ut hvor usannsynlig funksjonell sammenheng er for aminosyrer. Han startet med å lage en rekke varianter av et dårlig fungerende penicillin-inaktiverende enzym. I hver variant ble en sammenhengende klynge av 10 vedheng, byttet ut mot tilfeldige alternativer. Idéen var å finne ut hvor vanskelig det er for tilfeldige utvalg av vedheng å bli så funksjonelt koherente, som vedhengene de erstattet, noe som innebar at de akkurat var tilstrekkelig sammenhengende til at enzymet fungerte. Så snart dette var gjennomført for fire klynger, så var neste steg å kalkulere usannsynligheten ved at evolusjonære mekanismer skulle snuble borti slik minimale funksjonelle sammenhenger, ikke bare i disse 4, men alle de som trengs for at proteiner skal folde seg. Dette utførte Axe ved å omgjøre fraksjonen av mutanter som fungerte i de fire eksperimentene til en gjennomsnittlig sannsynlighet, av funksjonell sammenheng pr. aminosyre. Så multipliserte han disse for å estimere sannsynligheten til ett fullt randomisert gen som hadde denne funksjonelle sammenhengen, nødvendig for å forme en struktur, som støtter enzym-funksjon. Resultatet han fant var slående: Av de mulige gener som koder for protein-kjeder, på en lengde av 153 aminosyrer, så var det bare 1: 100 billioner billioner billioner billioner billioner billioner (10 opphøyd i 74), som er forventet å kode for en kjede som folder seg godt nok, til å utføre biologiske funksjoner. Dette tilsvarer sannsynligheten for blindt å finne ett hydrogen atom i ett verdensrom som er 28 milliarder lysår i diameter. Ett slikt lite målområde kan vi avskrive, som 'lost in space'. (5)
ville vært en lang kjede, i det å forme en stabil 3-dimensjonal struktur. Arrangert sekvensielt, på det viset, så passer vedhengene mer komfortabelt i deres foldede form, enn de ville gjort ved å flyte ustrukturert omkring i den cellulære væsken, slik som en tilfeldig sekvens av aminosyrer gjør. Uten deres fint-tilpassede formasjoner, kunne ikke livets proteiner utføre sine vitale funksjoner (4). I ett eksperiment prosjekt, satte Axe seg fore å finne ut hvor usannsynlig funksjonell sammenheng er for aminosyrer. Han startet med å lage en rekke varianter av et dårlig fungerende penicillin-inaktiverende enzym. I hver variant ble en sammenhengende klynge av 10 vedheng, byttet ut mot tilfeldige alternativer. Idéen var å finne ut hvor vanskelig det er for tilfeldige utvalg av vedheng å bli så funksjonelt koherente, som vedhengene de erstattet, noe som innebar at de akkurat var tilstrekkelig sammenhengende til at enzymet fungerte. Så snart dette var gjennomført for fire klynger, så var neste steg å kalkulere usannsynligheten ved at evolusjonære mekanismer skulle snuble borti slik minimale funksjonelle sammenhenger, ikke bare i disse 4, men alle de som trengs for at proteiner skal folde seg. Dette utførte Axe ved å omgjøre fraksjonen av mutanter som fungerte i de fire eksperimentene til en gjennomsnittlig sannsynlighet, av funksjonell sammenheng pr. aminosyre. Så multipliserte han disse for å estimere sannsynligheten til ett fullt randomisert gen som hadde denne funksjonelle sammenhengen, nødvendig for å forme en struktur, som støtter enzym-funksjon. Resultatet han fant var slående: Av de mulige gener som koder for protein-kjeder, på en lengde av 153 aminosyrer, så var det bare 1: 100 billioner billioner billioner billioner billioner billioner (10 opphøyd i 74), som er forventet å kode for en kjede som folder seg godt nok, til å utføre biologiske funksjoner. Dette tilsvarer sannsynligheten for blindt å finne ett hydrogen atom i ett verdensrom som er 28 milliarder lysår i diameter. Ett slikt lite målområde kan vi avskrive, som 'lost in space'. (5)
Fig 6. Rollen til amino-syre vedheng i å danne protein-strukturer
Oppfinnelser fra toppen-til-bunnen
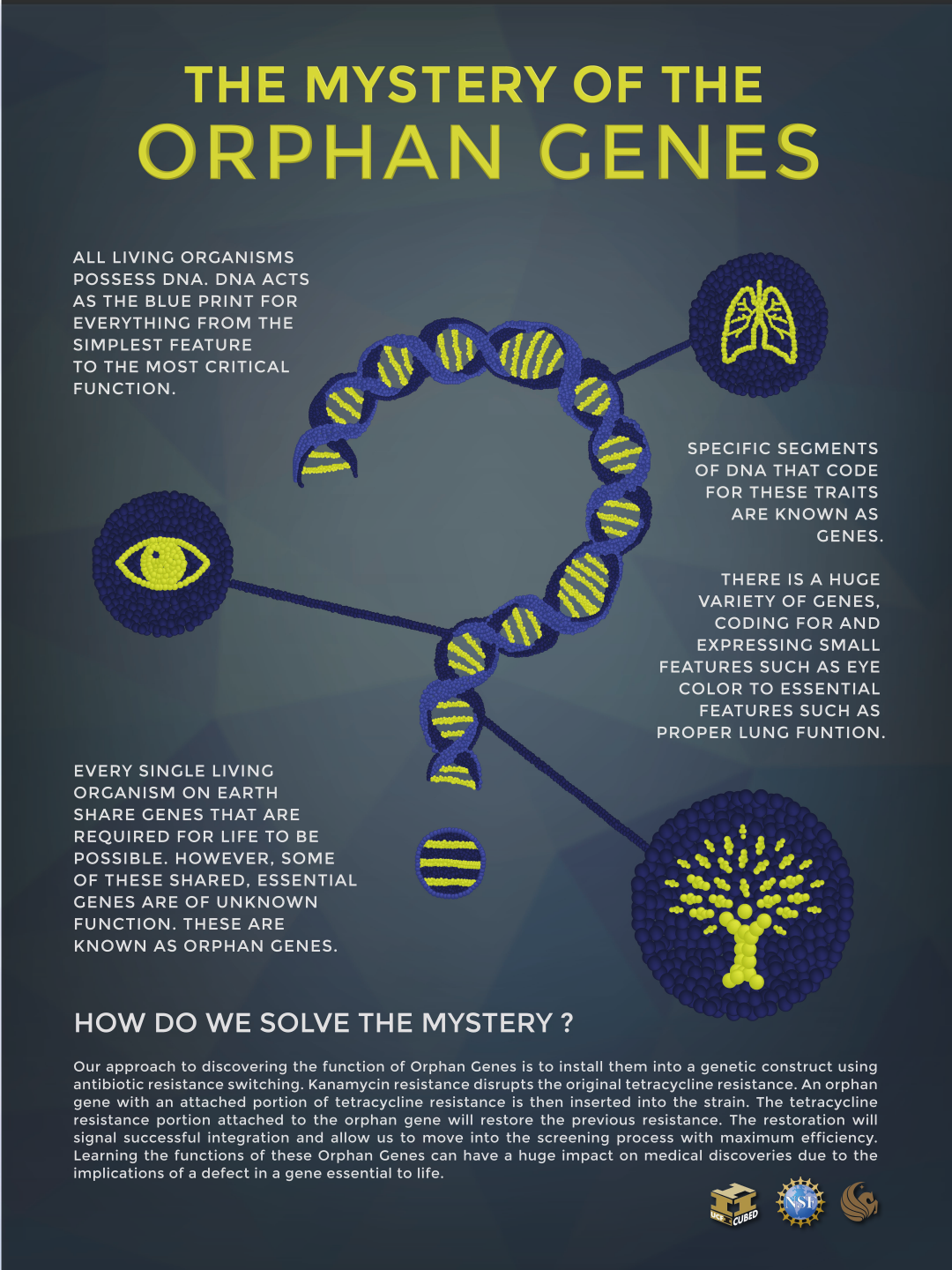 Så overbevist som Axe er over at protein-foldinger i seg selv er finurlige oppfinnelser, så ønsker han ikke å gi inntrykk av at alt av livets genistreker dreier seg om proteiner. Selvsagt gjør de ikke det. Som det skulle gå fram av Fig. 4 og 5, så er oppstillingen av aminosyrer bare étt aspekt ved livets eksplisitte design, og som også har en relativt lav posisjon i livets funksjonelle hierarki. Men mens oppfinnelsen av nye livsformer, utvilsomt er en høyere øvelse enn oppfinnelsen av nye protein-former, så synes den høyere øvelsen å kreve beherskelse av lavere-nivås øvelser. En av de store overraskelsene som kom fra genomprosjektet, er hvor mange unike gener og dermed proteiner som er tilstede i hver livsform, og inklusive former som for oss bare synes hårfint forskjellige. F.eks. undersøkte nylig en gruppe av tyske vitenskapsfolk genomsekvenser fra 16 cyanobakterielle strenger, i et forsøk på å skjelne alle de ulike gener disse strengene bar på (6). Siden alle var cyanobakterier, så kunne en tenke at de i hovedsak ville bære de samme gensettene, med små variasjoner. Forskerne fant at de delte ett felles sett på 660 gener, som er tilstrekkelig identiske til at de kan utøve samme biologiske funksjoner. Mer interessant var likevel deres funn av at nesten 14.000 gener, er unike for individuelle strenger. Av et gjennomsnitt på 869 unike gener pr streng, gjør dette bakterielle strenger mer ulike genetisk sett, enn like. Dette til tross for deres i hovedsak ytre likheter.
Så overbevist som Axe er over at protein-foldinger i seg selv er finurlige oppfinnelser, så ønsker han ikke å gi inntrykk av at alt av livets genistreker dreier seg om proteiner. Selvsagt gjør de ikke det. Som det skulle gå fram av Fig. 4 og 5, så er oppstillingen av aminosyrer bare étt aspekt ved livets eksplisitte design, og som også har en relativt lav posisjon i livets funksjonelle hierarki. Men mens oppfinnelsen av nye livsformer, utvilsomt er en høyere øvelse enn oppfinnelsen av nye protein-former, så synes den høyere øvelsen å kreve beherskelse av lavere-nivås øvelser. En av de store overraskelsene som kom fra genomprosjektet, er hvor mange unike gener og dermed proteiner som er tilstede i hver livsform, og inklusive former som for oss bare synes hårfint forskjellige. F.eks. undersøkte nylig en gruppe av tyske vitenskapsfolk genomsekvenser fra 16 cyanobakterielle strenger, i et forsøk på å skjelne alle de ulike gener disse strengene bar på (6). Siden alle var cyanobakterier, så kunne en tenke at de i hovedsak ville bære de samme gensettene, med små variasjoner. Forskerne fant at de delte ett felles sett på 660 gener, som er tilstrekkelig identiske til at de kan utøve samme biologiske funksjoner. Mer interessant var likevel deres funn av at nesten 14.000 gener, er unike for individuelle strenger. Av et gjennomsnitt på 869 unike gener pr streng, gjør dette bakterielle strenger mer ulike genetisk sett, enn like. Dette til tross for deres i hovedsak ytre likheter.
Forholdet av arts-spesifikke gener varierer fra en art til den neste, men deres eksistens i stort antall, synes å være en egenskap ved alt liv, ikke bare cyanobakterier. For å sitere sammendraget fra en teknisk artikkel nylig: "Sammenlignende genomanalyser indikerer at hver taksonomisk gruppe, så langt som det er undersøkt, inneholder 10-20% av gener som mangler gjenkjennelige homologe i andre arter." (7) M.a.o. så har alle arter mange gener som ved første øyekast synes å være enestående, ulik noen gener funnet andre steder. Det nøye arbeidet med å finne strukturen til proteinene som disse genene koder for, viser at omkring 2/3 viser seg å ligne tidligere kjente proteiner, mens den gjenværende tredelen forblir genuint ny. (8)
Bilde 3. ORFan-gener (Open Read Frames of unkown origin)
Opprinnelsen til nye kategorier av liv, synes dermed å kreve nye gener og proteiner. Av disse to så inneholder den ene det andre: akkurat som kyndighet i å lese og stave, bare er første steg mot skrivekyndighet, så er kyndighet i protein-design bare ett basis-steg, mot å mestre livets design. Faktumet at beherskelse av dette basis-steget fullstendig er utenfor rekkevidden til blinde evolusjonære mekanismer, innebærer evolusjonens fordervelse. Å tro på evolusjonsteorien innebærer sluttelig å tro noe som er mye mindre sannsynlig enn å gjentatte ganger treffe ett atom i vårt univers, ved 'blindt' å slippe sub-atomiske merker. Det skulle ikke noen tro.
Fruktene ved 'alminnelig vitenskap'
Det som vi har sluttet om oppfinnelser generelt, at de ikke inntreffer utilsiktet, er desto mer sant for de spesielt fabelaktige oppfinnelser vi ser i levende organismer. Vi kan slutte at protein-molekyler ikke dannes tilfeldig, og det som er tilfelle for dem, er enda mer tilfelle for høyere systemer som benytter fotosyntese og syn, og enda mer så for hele nivåer der mange funksjoner går sammen i én hensikt. Akkurat som dikt, instruksjoner og kjærlighetsdikt er fraværende i QWERTY-vrøvl, som kan finnes ved blinde søk, slikt er det med livet også. Hver ny livsform er dermed en mesteroppfinnelse, i sin egen rett, som bærer i seg din egen distinkte versjon av funksjonell sammenheng på høyeste nivå. Axe kan personlig bare se disse flinke, blomstrende, spinnende, pustende, gravende, flyvende, og kjempende oppfinnelser å komme fra Guds sinn. I følge Axe er det ikke noe annet som stemmer, og dermed blir det Gud vi har å takke for vår eksistens. At hver av livets arter har sin egen plass i vårt sinn, reflekterer det faktum at de ble gitt sin egen plass i verkstedet til det høyest rangerende sinn. Det er lite rom for kluss her, ingen tanke om at ett mesterverk flekker til et annet, som om briljante idéer kunne blandes lik maling. At vi kom fra det verkstedet skulle forbløffe oss desto mer. Av alle de undre som gjør jorda til sitt hjem, så er vi de eneste som stopper og reflekterer, og tar inn over oss hele dette oppsettet. Fem deler inspirerende og én del trøblete, og vet at intet av dette er ikke-tilsiktede tilfeldigheter. Barnas intuisjon viste seg riktig hele veien.
Bilde 4. Eks. på ødelagt design-intuisjon (Bilde brukt m. tillatelse)
 Referanser:
Referanser:
1. Thomas Nagel, "Mind and Cosmos: Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False (Oxford: Oxford University Pres, 2012), 32
2. For den teknisk innstilte anbefales denne oversikts artikkelen: I. Grotjohann and P.Fromme, "Structure of Cyanobacterial Photosystem I," Photosynthesis Research 85 (2005): 51-72
4. Termen 'i seg selv uordnede proteiner' refererer til en klasse proteiner som utnytter en delvis (eller fullstendig) ikke-foldet status, for å utføre visse biologiske funksjoner. Siden alt liv er avhengig av et stort antall av presist foldede proteiner, så er dette Axe sitt fokus. Argumentene og bevisene han fremfører for foldede proteiner affekteres ikke ved at det eksisterer proteiner som ikke folder seg.
5. Et suksessrikt søk etter ett foldet protein av denne størrelsen er ikke helt 'fantastisk' usannsynlig, derfor kan vi ikke si at det er fysisk umulig. Imidlertid har Michael Denton ett (1: 10 opphøyd i 40) kriterium, som tilsier at det er biologisk umulig. Dentons kriterim er mer relevant, siden det er snakk om et biologisk problem. Imidlertid gjør det liten forkjell (hvilket kriterium en bruker), fordi tilfeldige oppfinnelser som krever bare to nye proteiner, er umulige etter begge kriterier.
6. C. Beck et al., "The Diversity of Cyanobacterial Metabolism: Genome Analysis of Multiple Phototrophic Microorganisms," BMC Genomics 13 (2012): 56
7. K. Khalturin et al., "More than just Orphans: Are Taxinomically Restricted Genes Important in Evolution?" Trends in Genetics 25, no.9 (2009): 404-43
8. L. Jaroszewski, "Exploration of Uncharted Regions of the Protein Universe," PLoS Biology 7. no.9 (2009): e1000205
Stoffutvalg og bilder ved Asbjørn E. Lund